3.4 Visuelle Muster- und Objektbildung
| Zur Wahrnehmung von graphischen Elementen in kartographischen Präsentationen kann unterschieden werden, was vom Auge und Gehirn von den Elementen eigentlich aufgenommen und verarbeitet werden muss oder was eventuell gedanklich vorgegeben ist und was also „erinnert“ und „verglichen“ wird. | |
| Gustav Theodor Fechner (1860)Fechner, G.Th. (1860): Elemente der Psychophysik. 2 Bände, Leipzig hat u.a. zu dieser Frage die Begriffe distaler Reiz und proximaler Reiz geprägt, durch die die Reizeigenschaften der optischen Außenwelt (distaler Reiz) und die Reizverarbeitung durch die Sinnesorgane, also quasi das „Retinaabbild“ (proximaler Reiz), unterschieden, aber auch in einen ursächlichen Zusammenhang gestellt werden (vgl. Gundlach 1976 Gundlach, H. (1976): Reiz – zur Verwendung eines Begriffes in der Psychologie. Bern). Die Intension der visuellen Wahrnehmung liegt nach dieser Unterscheidung darin, dass durch die verschiedenen Prozesse der Reizverarbeitung ein möglichst dem distalen Reizkonstrukt angenähertes gedankliches Gebilde entstehen sollte. Auf der anderen Seite besteht aber im Wahrnehmungsprozess die Tendenz, dass bei starker Veränderung eines proximalen „Reizobjektes“ der Eindruck des Objekt erstaunlich konstant bleibt, die Außenwelt also mehr fluktuieren kann, als wahrgenommen wird (vgl. „Konstanzleistungen“, Kap. 2.5.5). | Distaler Reiz: Reizeigenschaften der optischen Außenwelt; Proximaler Reiz: Eigenschaften des „Retinaabbildes“; |
| Eine weitere Frage, die sich auch mit der Verarbeitung von visuellen Reizen beschäftig, ist die Überlegung, ob die Wahrnehmung auf der Basis von erfahrenen und erlernten Kategorien der realen Umwelt erfolgt oder ob nicht zusätzlich eine „biologisch zweckmäßige Konstruktion“ unseres Organismus zur zielorientierten und sinnvollen Aufnahme und Verarbeitung von Informationen beiträgt (vgl. Mausfeld 2005 Mausfeld, R. (2005): Wahrnehmungspsychologie: Geschichte und Ansätze In: Handwörterbuch Allgemeine Psychologie: Kognition. Göttingen). Wie Piaget und Inhelder Piaget, J. u. Inhelder, B. (1975a): Die Entwicklung des räumlichen Denkens beim Kinde. Gesammelte Werke, 6, Stuttgart eindringlich dargestellt haben (1975a Piaget, J. u. Inhelder, B. (1975a): Die Entwicklung des räumlichen Denkens beim Kinde. Gesammelte Werke, 6, Stuttgartu. 1975bPiaget, J. u. Inhelder, B. (1975b): Die natürliche Geometrie des Kindes. Gesammelte Werke, 7, Stuttgart), stehen gerade komplexe Wissens- und Fähigkeitsschemata zur räumlichen und geometrischen Wahrnehmung und gedanklichen Repräsentation schon im frühen Alter im umfangreichen Maße zur Verfügung und müssen daher wohl kaum erfahren oder erlernt werden, so dass das u.a. von Mausfeld erwähnte Konzept plausibel erscheint. | Frage zu Voraussetzungen von Wahrnehmungsprozessen: „erfahrene und erlernte Kategorien“ und/oder „biologisch zweckmäßige Konstruktion“ unseres Organismus? |
| Die Ziele der Wahrnehmung und der gedanklichen Verarbeitung kartographischer Informationen müssen vor diesem Hintergrund differenzierter betrachtet werden, als dies in der psychologischen und neurowissenschaftlichen Literatur meistens geschieht. Die dazu häufig abgehandelten Thematiken stellen besonders „Orientierungsweisen mit Karten“ in den Mittelpunkt der Betrachtung und dies nicht selten im Vergleich mit Vorgängen der „natürlichen Raumwahrnehmung“ und dem Agieren in der physischen Umwelt. | kartographischer Schwerpunkt in der Wahrnehmungsforschung sind Erkenntnisse zur „räumliche Orientierung in Karten“! |
| Dieser Gesichtspunkt der „Orientierung mit Karten“ ist sicherlich ein wichtiger Aspekt der Nutzung von so genannten Topographischen Karten oder beispielsweise des großen Repertoires an „Internet-Karten“, die zur Unterstützung von Bewegung oder zum Transport im Lebensraum geeignet sind bzw. die der visuellen Erschließung städtischer oder regionaler Räume mit Hilfe von konventionellen oder elektronischen Medien dienen. | |
| Ein Aspekt, der über diese Kartenfunktionen hinausgeht, betrifft Fragestellungen, die die fachlich spezialisierte Erschließung von georäumlichen Mustern, Sachverhalten und Prozessen z. T. unter Berücksichtigung von dreidimensionalen und zeitlichen Parametern zum Ziel haben. Die entsprechenden Thematiken werden meistens unter wissenschaftlichen Fragestellungen in so genannten Thematischen Karten erarbeitet, abgebildet und repräsentiert (vgl. Kap. 3.1). | Schwerpunkte in dieser Arbeit: Erkenntnisse zur “ fachlichen Erschließung von georäumlichen Mustern, Sachverhalten, Trends und Prozessen“. |

Abb. 34.1 Bedeutungsorientierte Informationsverarbeitung
(nach Bollmann 1977 Bollmann, J. (1977): Probleme der kartographischen Kommunikation. Bonn)
| Für die Einschätzung von Wahrnehmungsprozessen unterscheiden sich die beiden genannten Kartenfunktionen vor allem hinsichtlich des Ablaufs der visuell-kognitiven Herangehensweise an die Ableitung von Informationen und Wissen: So ist bei der „Orientierung mit Karten“ von einem kontinuierlichen Wechsel zwischen gezielter lage- und bedeutungsorientierter Identifizierung von Objekten (Zeichen) und von Objekten in deren Nachbarschaft auszugehen. | Orientierungsfunktion: lage- und bedeutungsorientierte Identifizierung von Objekten im Rahmen von Nachbarschaftsmustern. |
| Dies kann sich an Strecken, in Arealen oder auch an bestimmten ausgezeichneten Positionen von Orten unter Berücksichtigung von umgebenden regionalen Merkmalen und Eigenschaften vollziehen. Wie Abbildung 34.1 schematisch zeigt, ist das besondere Kennzeichen dieser Vorgänge der wiederholte visuell-gedankliche Rückgriff auf abgebildete Objekte bzw. auf Zeichenbedeutungen unter Berücksichtigung ihres räumlichen Verlaufs oder ihrer räumlichen Ausbreitung in Form eines merkfähigen „Wissenspfads“. | Vorgang der Orientierung: Wiederholter Rückgriff auf abgebildete Objekte unter Berücksichtigung ihrer räumlichen Ausbreitung und Umgebung als merkfähiger „Wissenspfad“. |
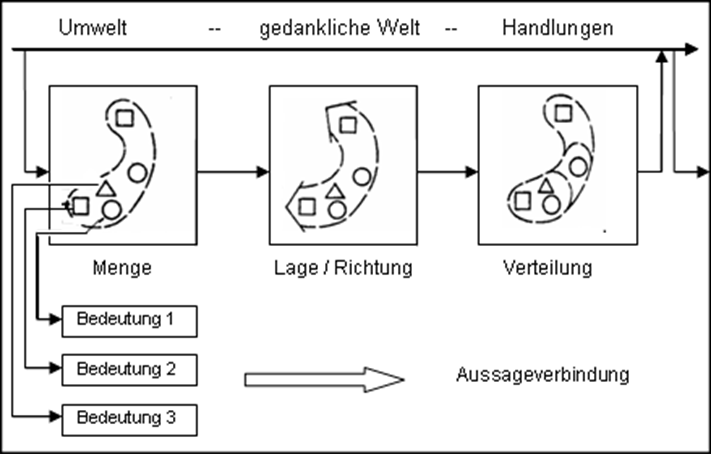
Abb. 34.2 Raumorientierte Informationsverarbeitung
(nach Bollmann 1977 Bollmann, J. (1977): Probleme der kartographischen Kommunikation. Bonn)
| Erheblich anders lässt sich der Wahrnehmungsablauf bei der Interpretation von Thematischen Karten vorstellen. Weniger bei der Identifizierung von einzelnen positionsgebundenen Objekten, Sachverhalten oder Bedeutungsmustern, als vielmehr bei der häufig im gesamten Gesichtsfeld (Kartenbild) erfolgenden Wissenserkundung (-exploration) ergibt sich u.a. ein mehrfaches und sich wiederholendes visuelles Absuchen nach Elementrelationen und Mustervarianten sowie nach deren Lage- und Richtungsausbreitung bzw. Verteilung. Dies führt zu georäumlichen Erkenntnissen über die analysierte Raumstruktur und zwar sowohl lokal unterschieden als auch zu ganzen Raumtypen aggregiert (vgl. die schematisierende Darstellung in Abbildung 34.2). | Vorgang der Wissensexploration: mehrfaches und sich verdichtendes Absuchen des Kartenbilds nach Elementrelationen und Mustervarianten sowie nach deren Lage- und Richtungsausbreitung bzw. Verteilungen. |
| In den weiteren Ausführungen steht diese zuletzt skizzierte Funktion von Karten im Mittelpunkt des Interesses (vgl. dazu auch die Ausführungen in Abschnitt B dieser Arbeit). Im Folgenden sollen hierzu als Einführung einige relevante Erkenntnisse aus der Psychologie und den Neurowissenschaften dargestellt werden. |
3.4.1 Neuronale Verarbeitung figurativer Reize
| Nach Gegenfurtner et al. (2010, S. 2) Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen kann zurzeit als relativ sichere neurowissenschaftliche Erkenntnisbasis von drei Zusammenhängen der elementaren visuellen Informationsverarbeitung ausgegangen werden (vgl. auch Bocianski 2010Bocianski, D.I. (2010): Lokalisation sukzessiv präsentierter Reize – Interaktion räumlicher Aufmerksamkeit und lokaler Verarbeitungsmechanismen, Diss., Philosophische Fakultät, RWTH Aachen): | Relativ sichere neurowissenschaftliche Erkenntnisbasis: |
|
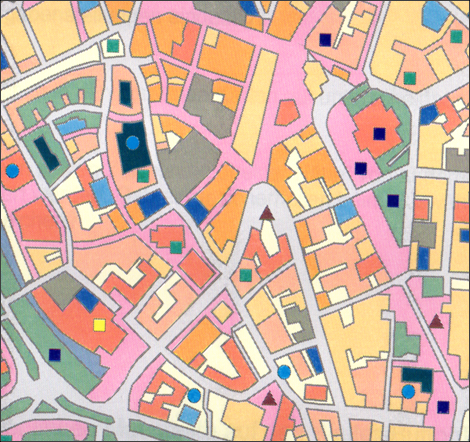

Abb. 34.3 Visuelle Stimulusstruktur am Beispiel einer Thematischen Karte
(aus Thoß 1996 Thoß, Ch. (1996): Empirische Untersuchung zur Informationsverarbeitung im kartographischen Wahrnehmungsraum „Problemraum“. Unveröfft. Studienarbeit, Universität Trier)
| Bei Betrachtung des in Abbildung 34.3 präsentierten Ausschnitts einer Testvorlage aus einem Forschungsprojekt der Empirischen Kartographie der Universitär Trier (Thoß 1996 Thoß, Ch. (1996): Empirische Untersuchung zur Informationsverarbeitung im kartographischen Wahrnehmungsraum „Problemraum“. Unveröfft. Studienarbeit, Universität Trier) wird deutlich, welche anspruchsvollen elementaren visuellen Leistungen erforderlich sind, um die optisch angebotenen graphischen Elemente und Musterstrukturen visuell unterscheiden zu können und darüber hinaus deren Formen, Größen, Farben, Helligkeitsabstufungen und Umrandungen lagemäßig abzugrenzen bzw. zu verbinden sowie kognitiv zu ordnen und gedanklich in merkfähige Konstrukte zu überführen. Es ist davon auszugehen, dass diese Leistungen aus komplexen sensorischen und kognitiven Prozessen resultieren, deren psychologischen und neurowissenschaftlichen Deutungen im Zusammenhang mit den oben bei Gegenfurtner aufgeführten Zusammenhängen der visuellen Informationsverarbeitung auf dem momentanen Stand sicherlich noch nicht abgeschlossen sind. | Zur Identifizierung und Einordnung von optisch angebotenen graphischen Elementen und Musterstrukturen in Karten sind komplexe sensorische und kognitive Prozesse erforderlich. |
| Gegenfurtner Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen führt aus, dass „gegenwärtig nur unser Wissen über die ersten Stufen der visuellen Verarbeitung ausgehend von der Retina bis zur primären Sehrinde, V1, relativ groß ist. So wissen wir zum Beispiel wie die visuelle Information in V1 verteilt wird und welche Eigenschaften die Neurone in den einzelnen Schichten von V1 besitzen. Von den sich anschließenden höheren sekundären und tertiären Verarbeitungsebenen ist zwar bekannt, auf welche visuellen Reize (….) einzelne Neurone maximal antworten, aber wie es zu dieser erstaunlichen Selektivität kommt, ist weitgehend unklar“ (2010, S.2 Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen). | Über die erste Verarbeitungsstufe in der Retina und primären Sehrinde des Gehirns ist relativ viel, über Vorgänge in höheren sekundären und tertiären Verarbeitungsebenen ist neurowissenschaftlich eher weniger bekannt. |
| Die bildgetreue Repräsentation von Mustern durch Weiterleitung graphischer, euklidischer, topologischer und dreidimensionalen Relationen kann von den Eingangsrezeptoren im Auge bis in die höheren Ebenen im Gehirn verfolgt werden, auch wenn die Feststellung von Relationen in höheren Regionen des Gehirns unsicherer wird. So ist weiterhin die Bildung von funktionalen Netzwerken visueller Neuronen, denen bestimmte Aufgaben und Funktionen zu kommen, möglich. Dabei lassen sich die Merkmalunterschiede zwischen den einzelnen Cluster zum Teil deutlich feststellen. Die Unterschiede betreffen die Form der eintreffenden Signale, die neuronalen Reaktionen und ihre Verschaltungen. Außerdem kann eine zunehmende Aufgaben- und Funktionsverteilung entlang der neuronalen Verarbeitungsbahnen, die in Abhängigkeit von der Komplexität der visuellen Reize eine maximale neuronale Antwort hervorrufen, nachvollzogen werden. So gibt es in der primären Sehrinde neuronale Antworten auf elementare visuelle Elemente und dagegen im unteren Temporallappen nur relativ einfache Reaktionen auf komplexe visuelle Strukturen. Hinzu kommen in den höheren Verarbeitungsebenen Beeinflussungen durch Faktoren wie Aufmerksamkeit, Gedächtnis oder mit dem Reiz verbundenen Handlungen. | Die bildgetreue Repräsentation von Mustern durch Weiterleitung graphischer, euklidischer, topologischer und dreidimensionalen Relationen kann von den Eingangsrezeptoren im Auge bis in die höheren Ebenen des Gehirns verfolgt werden. |
| Im Folgenden werden drei Bereiche herausgehoben, die für elementare Prozesse der Wahrnehmung kartographischer Elemente relevant sind und die auch in entsprechender neurowissenschaftlichen Literatur häufig im Zentrum der Betrachtung stehen. | In der Literatur stehen drei Bereiche im Zentrum der Betrachtung: |

Abb. 34.4 Machsche Streifen
3.4.1.1 Abgrenzung graphischer Elemente
| Die in Abbildung 34.3 wirkenden flächigen Elemente sind zum einen aufgrund ihrer Farbton- (Grauton-) Ausprägungen von Interesse, zum anderen aber auch aufgrund ihrer Abgrenzungen zueinander. Aus diesen Abgrenzungen können u.a. Unterschiede, Identitäten, Umrissformen und Lagerelationen als relevante Informationen extrahiert werden. In der Präsentation sind zur visuellen „Sicherheit“ die einzelnen Elemente durch graue Umrandungen von ihren unmittelbaren Nachbarn abgegrenzt. Bei „deutlichen“ Farbtonunterscheidungen erscheint dies zur Farbstrukturierung eher überflüssig bzw. „redundant“ zu sein (vgl. Bollmann 1977, S. 128 Bollmann, J. (1977): Probleme der kartographischen Kommunikation. Bonn), bei geringen Farbtonunterschieden oder bei kleinen Flächen sind die Umrandungen dagegen eher hilfreich, um eine sichere Identifizierung und Unterscheidung zu gewährleisten. | Aus Abgrenzungen können Unterschiede, Identitäten, Umrissformen und Lagerelationen als relevante Informationen extrahiert werden. |
| Bei der Wahrnehmung in der natürlichen Umwelt oder von Fotographien und Satellitenbildern fehlen diese „unterstützenden“ optischen Begrenzungslinien, so dass beim Auftreten geringer Farb- oder Helligkeitsunterschiede es eventuell zu „Verwischungen“ oder „Nivellierungen“ kommen kann, mit der Folge, dass Grenzbereiche von Elementeinheiten nicht eindeutig identifiziert werden. Auf dieser Abgrenzungsmöglichkeit basiert aber im Wesentlichen die visuelle Strukturierung der wahrgenommen Realität. Das visuelle System stellt dazu einen neuronalen Mechanismus bereit, der den Eindruck dieser Abgrenzung verstärkt. Ein bekanntes Beispiel für die Demonstration dieses Mechanismus sind die in Abbildung 34.4 präsentierten „Machsche Streifen“ (vgl. Mach 1865 Mach, E. (1865): Über die Wirkung der räumlichen Vertheilung des Lichtreizes auf die Netzhaut. Sitzungsberichte der mathematisch-naturwissenschaftlichen Classe der kaiserlichen Akademie der Wissenschaften, 52, 303–322), bei denen gezeigt wird, dass bei unterschiedlich grauen Streifen die Grenzbereiche zwischen den Grautönen jeweils aufgehellt bzw. abgedunkelt, also kontrastreicher erscheinen. | Machsche Streifen demonstrieren, dass die Grenzbereiche bei aneinanderstoßenden, unterschiedlich grauen Flächen kontrastreicher erscheinen. |
| Die Ursachen für diese außerordentlich wichtigen visuellen Leistungen liegen im neuronalen Mechanismus der lateralen Inhibitation begründet, dessen Entdeckung und Formulierung vor allem auf Stephan William Kuffler (1953)Kuffler, S.W. (1953): Discharge patterns and functional organization of the mammalian retina. Journal of Neurophysiology, 16 zurückgeht. Wie in Kapitel 1.2.5 und an anderen Stellen schon ausgeführt wurde, handelt es sich um einen Mechanismus, bei dem zwischen zwei oder mehr Nervenzellen sogenannte Interneuronen geschaltet sind, wodurch erregte Zellen von benachbarten Nervenzellen wieder um einen gewissen Betrag gehemmt werden. Bei diesen Interneuronen handelt es sich um Horizontalzellen der Retina, die nur bei der Verschaltung von Zapfen eine Rolle spielen. |
Durch den Mechanismus der lateralen Inhibitation werden erregte Zellen von benachbarten Nervenzellen wieder um einen gewissen Betrag gehemmt. |
| Zur Verdeutlichung der bei der lateralen Hemmung wirkenden Bedingungen lässt sich der Gesamtzusammenhang der Mechanismen folgendermaßen darstellen: | Gesamtzusammenhang der lateralen Inhibitation: |
|
|
| Mit Unterstützung dieses visuellen Effektes können die im Gesichtsfeld angebotenen optischen Strukturen verstärkt in ihren wirkenden Reizverteilungen und -ausprägungen vor allem in ihren Grenzbereichen unterschieden und dadurch in ihren Ausbreitungen und formenhaften Verläufen visuell strukturiert werden. | Durch diesen visuellen Effekt werden die im Gesichtsfeld angebotenen optischen Strukturen in ihrem Unterschied verstärkt. |
3.4.1.2 Bildung Neuronaler Muster (Retinotopie)
| Die visuell-kognitive Verarbeitung graphischen Szenerien hat das Ziel, optische Reize strukturiert zu enkodieren und bestimmte Ausschnitte gedanklich zu repräsentieren. Ziel kann sein, relevante Merkmale unmittelbar mit anderen Szenerien zu vergleichen oder ganze bildliche Ansichten im Gedächtnis zu behalten und diesen Gedächtnisbesitz bei Bedarf in „Anwesenheit“ der ursprünglichen Reize wieder zuerkennen (engl. rekognition) oder ohne präsentierten Reize wieder zuerinnern (engl. recall). Nach Schumann-Hengsteler (1995, S. 29ff) Schumann-Hengsteler, R. (1995): Die Entwicklung des visuell-räumlichen Gedächtnisses. Göttingen sind beide Gedächtnisleistungen sowohl kurzfristig (Kurzzeitgedächtnis) als auch längerfristig (Langzeitgedächtnis) möglich und können z.B. im georäumlichen Handlungsprozess bei der Transformation von medial geprägten Informationen in problemorientiertes Wissen eine wichtige Rolle spielen (vgl. Bollmann et al. 2000Bollmann, J. u. Uthe, A.-D. (2000): Kartographische Steuerung georäumlicher Erkenntnisprozesse. In: Geowissenschaftliche Mitteilungen der Technischen Universität Wien. 53, 31-47, Wien.). Gerade bei der Verarbeitung bildlicher Informationen sind diese genannten Prozesse besonders anspruchsvoll und daher auch ein relevantes Thema der psychologischen und neurowissenschaftlichen Forschung. |
Die Informationsverarbeitung in Karten kann das Ziel haben, Merkmale mit anderen Szenerien zu vergleichen oder bildliche Ansichten im Gedächtnis zu behalten und bei Bedarf in „Anwesenheit“ der ursprünglichen Reize wieder zuerkennen oder ohne präsentierte Reize wieder zu erinnern. |
| Eine weit reichende Eigenschaft des Gehirns unterstützt dabei die effektive Verarbeitung haptisch, visuell und akustisch ermittelter Informationen. Beispielweise erfolgt bei haptischen Informationen (haptisch = den Tastsinn betreffend, „fühlbar“; vgl. z.B. Grunwald 2011 Grunwald, M. (2011): Das Sinnessystem Haut und sein Beitrag zur Körper- Grenzenerfahrung. In: Körperkontakt. Multidisziplinäre Erkundungen. 29–54, Giessen) durch den somatosensorische Cortex, als Teil der Großhirnrinde, eine Abbildung des zentralen Bereichs der durch verschiedene Körperabschnitte ermittelten Reize. Dieser Bereich des Gehirns ist streng somatotop aufgebaut, das bedeutet, dass haptische Informationen, die in benachbarten Zonen des Körpers aufgenommen werden, auch in entsprechend nebeneinander liegenden Arealen der Cortex organisiert sind. So entsteht eine Abbildung von Körperzonen auf der Hirnrinde, allerdings entsprechend der jeweiligen Wertigkeit verzerrt. Beispielsweise werden haptische Empfindungen, die durch Hand, Finger, Gesicht und Lippen entstehen, zwar in benachbarten Arealen verarbeitet, sind aber – wahrscheinlich entsprechend ihres relativ hohen haptischen Stellenwertes – gegenüber anderen Körperregionen besonders verbreitet angelegt,. Die Übertragung der Anordnung von Körperarealen in entsprechend verteilten Hirnregionen wird als Somatotopie und deren Darstellung als somatope Karten bezeichnet (vgl. z.B. Birbaumer et al. 2010 Birbaumer, N. u. Schmidt, R.F. (2010): Biologische Psychologie. Berlin, Ruben 2011 Ruben, J. (2011): Somatotope Organisation des sekundären somatosensorischen Kortex des Menschen. Diss., Medizinischen Fakultät Charité – Universitätsmedizin Berlin). | Eine weit reichende Eigenschaft des Gehirns unterstützt die effektive Verarbeitung ermittelter Informationen, indem Informationen, die in benachbarten Zonen des Körpers aufgenommen werden, auch in entsprechend nebeneinander liegenden Arealen der Cortex organisiert sind. |
| Interessant ist diese haptische Wahrnehmung auch im Zusammenhang mit sog. „taktilen Schnittstellen“ im Sinn der Virtual Reality, bei denen ein fühlbares Feedback von Ausgabegeräten an den Benutzer in Form von Krafteinwirkungen angestrebt wird. Ziel ist die möglichst realistische Empfindung einer simulierten Umwelt (Müller 2002 Müller, A. (2002): Virtual Reality Modeling Language. In: Lexikon der Kartographie und Geomatik. 2, Heidelberg). | |
| Für die visuelle Wahrnehmung ist dieses Organisationsprinzip zwischen Reizaufnahme und mentaler Weiterverarbeitung noch umfassender realisiert. So bleibt die topologische Struktur von Orten auf der Retina mit bestimmten Zellerregungen bei der neuronalen Verarbeitung von Informationen über den Corpus geniculatum laterale, CGL (vgl. Kapitel 2.5.3) im primären visuellen Kortex erhalten. Das bedeutet, dass die räumlich benachbarte Anordnung von lichtempfindlichen Zellen auf der Retina als lagemäßig ähnlich liegende Zellen im Gehirn abgebildet wird (vgl. z.B. Hoffmann et al. 2010 Hoffman, D. D. u. Richards, W. (1985): Parts of Recognition. Cognition, 18, 65-96). | Das bedeutet für die visuelle Wahrnehmung, dass die räumlich benachbarte Anordnung von lichtempfindlichen Zellen auf der Retina als lagemäßig ähnlich liegende Zellen im Gehirn abgebildet wird. |
| Bei Gegenfurtner et al. (2010) Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen ist der Zusammenhang der neuronalen Mechanismen in den verschiedenen Bereichen, Arealen und Schichten der Retina und des Gehirns sinngemäß folgendermaßen zusammengefasst: In der Retina reagieren Ganglienzellen auf Reize, die Photorezeptoren in einem bestimmten Bereich des visuellen Feldes erregt haben. Diesen Bereichen des Gesichtsfeldes entsprechen rezeptive Felder von Neuronen (vgl. Kapitel 1.2.2), sie lassen sich relativ genau in ihren Positionen auf der Retina bestimmen. So besitzen nebeneinander liegende retinale Ganglienzellen gleichfalls benachbarte und sich überlappende rezeptive Felder und projizieren zu benachbarten Neuronen der nächsthöheren Stufe. Diese räumliche Ordnung bleibt von der Rezeptorebene in der Retina bis in die höheren Verarbeitungsebenen im Kortex weitgehend erhalten, auch wenn die rezeptiven Felder von Stufe zu Stufe zunehmend größer werden. | So besitzen nebeneinander liegende retinale Ganglienzellen gleichfalls benachbarte und sich überlappende rezeptive Felder und projizieren zu benachbarten Neuronen der nächsthöheren Stufe. |
| „Durch den Erhalt der räumlichen Anordnung der retinalen Ganglienzellen bei der Projektion sind auch die Geniculatumschichten retinotop organisiert. Die sechs Schichten sind also so angeordnet, dass die sechs neuronalen Karten der kontralateralen Gesichtsfeldhälfte genau übereinander liegen, und sich daher auch die Zentren der rezeptiven Felder von vertikal übereinander liegenden Neuronen aller Schichten an derselben Stelle befinden. Das Geniculatum dient nicht nur als Durchgangsstation für die Eingangssignale vom Auge zum primären visuellen Kortex, sondern es erhält auch zahlreiche Eingänge aus dem Kortex oder vom Hirnstamm“ (Gegenfurtner et al. 2010. S.3 Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen) | Beim seitlichen Kniehöcker des Zwischenhirns (Geniculatum – CGL) liegen die sechs neuronalen Karten der kontralateralen Gesichtsfeldhälfte genau übereinander, so dass sich auch die Zentren der rezeptiven Felder von vertikal übereinander liegenden Neuronen aller Schichten an derselben Stelle befinden. |
| Wie oben angedeutet wurde, ergeben sich bei der lokalen Abbildung evozierter Potentiale im primären visuellen Kortex zwar topologische Übereinstimmungen mit dem Retinaabbild, es treten aber systematische euklidische Verzerrungen – als so genannte foveale Vergrößerung – vor allem im zentralen Gesichtsfeld auf, die zuerst von Tootell et al. (1982) Tootell, R.B.; Silverman, M.S.; Switkes E. u. de Valois R. L. (1982): Deoxyglucose analysis of retinotopic organization in primate striate cortex. Science 218, 902-904 untersucht und vermessen worden sind. | Bei den topologischen Übereinstimmungen mit dem Retinaabbild treten systematische euklidische Verzerrungen – als so genannte foveale Vergrößerungen – auf. |
| Diese verzerrte retinotope Repräsentation gilt nicht nur für den primären visuellen Kortex, sondern tritt auch in höheren visuellen Arealen auf. Der Grund liegt vermutlich darin, dass die Anzahl von retinalen Zellen im Bereich der Fovea und damit die Dichte der rezeptiven Felder weitaus höher bzw. größer sind als in der Peripherie der Retina und damit zu ihrer Repräsentation einen größerer Zellbereich erforderlich ist. Dadurch sind bei Aufnahmen mit Hilfe bildgebender Verfahren, wie z.B. der „funktionellen Magnetresonanztomografie“ (fMRT) (vgl. Hoffmann et al. 2010 Hoffmann, M.B.; Kaule, F.; Grzeschik, R.; Behrens-Baumann, W. u. Wolynski, B. (2010)), in der retinotopen Karte diese Bereiche größer abgebildet. Mit der fMRT-Technik können nicht invasive Gehirnprozesse im lebendigen Organismus (in vivo) untersucht werden. Dabei ist sowohl die zeitliche als auch die räumliche Auflösung deutlich niedriger als bei Einzelzellableitungen – es kann dadurch das Gehirn aber als Einheit untersucht werden (vgl. Schira 2004 Schira, M.M. (2004): „Die Verarbeitung von Konturen im visuellen Kortex des Menschen.“. Diss., FB Biologie/Chemie, Univ. Bremen). Untersuchungen zum Grad dieser Verzerrungen sowie ihre Darstellung oder Berechnung, z.B. mit Hilfe von „Verzerrungstensoren“ (vgl. z.B. Altenbach 2012 Altenbach, H. (2012): Kontinuumsmechanik. Berlin), führen zu einer Beschreibung der Position und Ausbreitung des u.a. durch Krümmung erzeugten Überschuss an kortikaler Fläche aus Sicht der Netzhaut. | Der Grund liegt vermutlich darin, dass die Dichte der rezeptiven Felder im Bereich der Fovea weitaus größer ist als in der Peripherie der Retina und damit zu ihrer Repräsentation ein größerer Zellbereich erforderlich ist. |
| Diese Aufnahmen und Berechnungen sind die Voraussetzung, um Detailanalysen zu einer Vielzahl visueller Areale vornehmen und beispielsweise Fragestellungen zur Entwicklungsplastizität des Gehirns untersuchen zu können (vgl. Dudel et al. 2001 Dudel, J.; Menzel, R. u. Schmidt, R.F., Hrsg. (2001): Neurowissenschaft: Vom Molekül zur Kognition. Berlin). Zu dieser sog. neuronalen Plastizität wird vermutet, dass sich z.B. Eigenschaften von Synapsen, Nervenzellen oder auch ganzen Hirnarealen in Abhängigkeit von der Verwendung in ihren Eigenschaften verändern bzw. anpassen (vgl. z. B. Doidge 2008 Doidge, N. (2008): Neustart im Kopf: wie sich unser Gehirn selbst repariert. Frankfurt a. M. u. New York). Dies könnte auch dazu führen, dass sich im Bereich der visuellen Wahrnehmung durch mentale Anpassungen neue oder noch unbekannte visuelle Szenerien besser verarbeiten lassen (vgl. vgl. Abschnitt B dieser Arbeit). | Die Aufnahmen und Berechnungen dieser Phänomene sind die Voraussetzung, um Fragestellungen zur Entwicklungsplastizität des Gehirns untersuchen zu können. |
3.4.1.3 Trennung optischer Signale
| Zur Erläuterung von visuellen Leistungen, die vom Organismus bei der Wahrnehmung von kartographischen Präsentationen erbracht werden müssen, ist erstens die visuelle Kontrastbildung zur Hervorhebung von Konturen erläutert worden. Zweitens wurden Mechanismen bei sog. retinotoper Karten als topologisch-euklidische Abbildung visueller Reize und Signale in der Retina und in Arealen des Gehirns beschrieben. Diese Mechanismen führen vermutlich zur effektiveren Verarbeitung visueller Informationen. | |
| In einem dritter Bereich wird nun skizziert, wie diese Verarbeitung so ungestört wie möglich erfolgen kann, dass z.B. die daraus abgeleiteten Informationen so verarbeitet werden, dass sie z. B. zielorientiert in Handlungen integriert werden können. Dazu verfügt das Wahrnehmungssystem über neuronale Einrichtungen, mit deren Hilfe bestimmte Arten von Reize getrennt und deren Aktionspotentiale über spezialisierte Übertragungskanäle an das Gehirn weiter geleitet und dort in höheren Arealen wieder gemeinsam verarbeitet werden. Von D. Hubel und T. Wiesel (1959 Hubel, D.H. u. Wiesel T.N. (1959): Receptive fields of single neurons in the cat`s striate Cortex. J. Physiol. 148, 574-591, 1962 Hubel, D.H. u. Wiesel, T.N. (1962): Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J Physiol, 160, 106-154) wurde dazu zuerst das Antwortverhalten von Neuronen im primären visuellen Kortexareal V1 auf visuelle Reize untersucht. | Das Wahrnehmungssystem verfügt über neuronale Einrichtungen, mit deren Hilfe bestimmte Arten von Reize getrennt und deren Aktionspotentiale über spezialisierte Übertragungskanäle an das Gehirn weiter geleitet werden. |
| Anders als die Neurone in der Retina oder im Geniculatum antworteten V1-Neurone nur schwach oder gar nicht auf punktförmige Lichtreize, aber sehr heftig auf kurze Lichtstreifen. Je nach Art des visuellen Reizes, der die größte Antwort hervorrief, werden drei unterschiedliche Zellentypen unterschieden (Hubel et al. 1962 Hubel, D.H. u. Wiesel, T.N. (1962): Receptive fields, binocular interaction and functional architecture in the cat’s visual cortex. J Physiol, 160, 106-154, Gegenfurtner et al. 2010 Gegenfurtner, K.R.; Walter, S. u. Braun, D.I. (2010): Visuelle Informationsverarbeitung im Gehirn. Allgemeine Psychologie, Justus-Liebig-Universität, Gießen und Sczepek 2011, S.32ffSczepek, J. (2011): Visuelle Wahrnehmung. Norderstedt): |
V1-Neurone antworten nur schwach oder gar nicht auf punktförmige Lichtreize, aber sehr heftig auf kurze Lichtstreifen. Es gibt also Zelltypen, die unterschiedlich auf unterschiedliche visuelle Reize reagieren.
|
|
|
| Die Zellen werden im Verlauf der Verarbeitung also immer spezialisierter gesehen und sprechen auf immer größere Bereiche des visuellen Feldes an. So werden in einem hierarchisch abgestimmten Prozessverlauf aus Kanten, deren Richtungen und Bewegungen sowie verteilten Ecken und Winkel komplexe Gebilde konstruiert, die der Erscheinung von Objekten der Umwelt entsprechen. Das heißt, es wird deutlich, dass es sich dabei um einen aktiven Prozess der Informationsverarbeitung handelt und nicht um einen statischen Vorgang, bei dem graphische Präsentationen als „einheitliches Bild“ verarbeitet werden. | Die Zellen werden im Verlauf der Verarbeitung also immer spezialisierter gesehen und sprechen auf immer größere Bereiche des visuellen Feldes an. |